
.
Au cours de l’ère Secondaire, alors que dominaient gymnospermes et reptiles, apparurent timidement les angiospermes. Chez ces plantes, le siège de la sexualité est un édifice complexe et hautement performant : la fleur. Elle se transforme en un fruit, produit par la plante-mère, qui contient les graines enfermées : l’embryon est doublement protégé. Dans le même temps, apparaissent les premiers mammifères, dont les jeunes étaient pris en charge et nourris par la mère.
.
La fin de l’ère Secondaire fut brutale, marquée par une phase de cataclysmes : les forêts de conifères géants et le peuplement reptilien furent anéantis par la sécheresse et le froid (peut-être à la suite de l’impact d’une grosse météorite ?) auxquels ne réchappèrent que quelques survivants dont la descendance nous est parvenue.
.
Les plantes à fleurs (elles sont à présent partout dominantes) sont aussi constituées d’une succession d’articles, qui comportent chacun une tige, une feuille et un bourgeon, à l’aisselle des deux autres. Les feuilles peuvent être alternes, opposées ou verticillées (à partir de trois au même niveau).
.
.
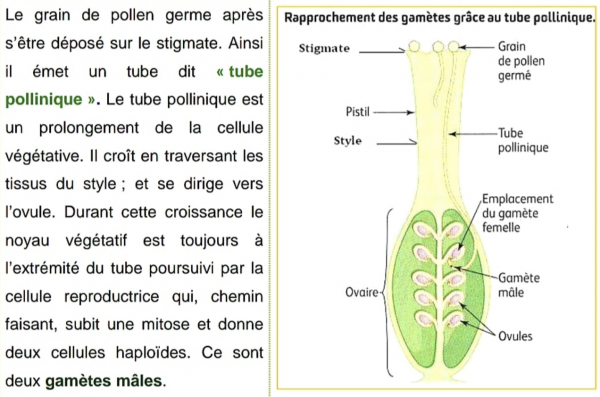
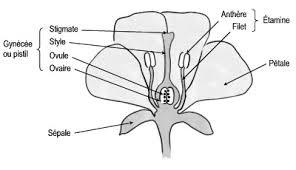
Dans les plantes à fleurs, l’article terminal se transforme, les organes reproducteurs se groupent en fleurs. Les feuilles sous la fleur s’appellent les bractées, puis les sépales, puis les pétales. Enfin, l’une d’elle se retourne pour donner le pistil, qui conduira à l’ovaire, d’autres se différencient en étamines, sacs à pollen (grains inertes).
.
Les étamines sont les organes mâles au sein desquelles s’opère la méiose. De cette méiose va résulter les grains de pollen, éléments mâles fécondants haploïde, cellules non cloisonnées (sincytium) qui contiennent deux noyaux qui sont libérés et quand ils atteignent l’ovule, il y a une nouvelle division : des deux spermatozoïdes, l’un fécondera l’ovule (comme pour les groupes précédents), l’autre s’unit avec deux des noyaux polaires du prothalle femelle, pour donner un massif cellulaire à « 3 n » chromosomes, l’alumen, qui envahit la cavité de l’ovule et se substitue au prothalle femelle dans son rôle nourricier. Cette « double fécondation » (découverte en 1899) permet une économie, car ce riche tissu de réserve (polyploïde) n’est mis en place que si la fécondation a eu lieu.
.

.
Après la fécondation (fusion des gamètes à l’intérieur de l’ovaire), les graines se développent et donnent des embryons en vie latente. Ils sont organisés, mais leur développement est stoppé jusqu’à ce que les conditions soient favorables. Ceci constitue aussi un progrès considérable car tous les végétaux inférieurs gaspillent leurs embryons. Pour toutes ces raisons, les plantes à fleurs sont dites « vivipares ». L’évolution vers une meilleure protection des embryons a poussé ces plantes à les enfouir toujours plus profond : on distingue ainsi les ovaires supères (archaïques) et infères (évolués). Cette sophistication est à comparer avec la meilleure protection assurée aux embryons des mammifères chez les placentaires que chez les marsupiaux (qui furent rapidement dominés, sauf dans quelques régions isolées du monde : Madagascar, Australie …).
.
Après la fécondation, l’ovaire se transforme donc en un fruit qui peut être :
–> déhiscent (ou sec), les plus primitifs, libération de nombreuses graines souvent pourvues d’un dispositif favorisant le transport par l’air (aigrette plumeuse) ou les animaux (crochets),
–> une baie, fruit secondairement charnu, dans un but de dissémination par les animaux (ex.: le Gui et les oiseaux),
–> indéhiscent (les graines restent dans le fruit), akènes (secs) ou drupes = fruit charnu à noyau (ex.: la cerise, l’olive).
.
Les plantes à fleurs sont essentiellement hermaphrodites, car pistil et étamines sont réunis dans la même fleur. Certaines au contraire sont dioïques (ex.: saules, kiwi, mercuriale, orties), c’est à dire qu’il existe des pieds mâles et des pieds femelles, bien évidemment, seuls les individus femelles donnent des fruits, ou monoïques, c’est à dire que des fleurs des deux sexes se développent sur le même arbre mais séparément (cas des arbres à chatons : chênes, peupliers, châtaigniers …). Certaines (comme le houx) avortent au moment de la floraison une partie de ses fleurs : elle est mâle ou femelle selon les saisons ! Dans le cas du Papayer, les deux arbres sont assez différents, mais l’ambiguïté persiste, car à partir d’un papayer mâle, on peut obtenir un papayer femelle en augmentant notablement les apports en eau et en engrais azoté.
.
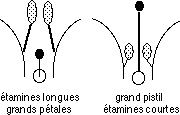
Lorsque le pollen d’une fleur féconde la graine de la même fleur, il y a brassage génétique (deux méioses distinctes), même si le stock reste le même. Dans le cas où le pollen provient d’un individu et féconde un autre individu végétal, le fonctionnement de la sexualité est optimal, le renouvellement génétique plus complet d’une génération à l’autre. C’est pourquoi la plupart des plantes à fleurs utilisent des modalités de rencontre pollen/graine permettant une fécondation croisée. Dans le cas des Campanulacées (ex.: Lobelia), la jeune fleur montre d’abord des étamines mûres libérant leur pollen, puis 48 heures après, le pistil s’épanouit et devient réceptif. Il y a disjonction des sexes dans le temps. Dans d’autres cas, il y a disjonction dans l’espace : le même individu porte des fleurs mâles et femelles séparées (arbres à chatons).
.
.
On observe aussi d’autres types de différenciation sexuelle chez les végétaux, la différenciation morphologique. C’est une découverte de Darwin sur la fleur de Primevère (Primula) : les deux types d’individus, fonctionnellement hermaphrodites, ne peuvent se féconder que sur l’autre type de fleur, ce n’est pas un sexe gamétique (génotypique), mais au niveau des individus dans une même population (sorte de sexe phénotypique). Chaque pied de primevère donnera 50% de descendance de chacune des deux formes ! Ce phénomène s’est révélé assez répandu.
.
La notion de sexe s’élargit encore avec des labiées (comme le Romarin et la Lavande) et certaines fabacées. Toutes les fleurs sont hermaphrodites, mais la fécondation n’est possible qu’entre deux individus différents. Il y a toujours deux sexes gamétiques, mais autant de sexes phénotypiques (dont la rencontre est fertile) que d’individus. Le support de ce phénomène d’exclusion est lié à la recon-naissance de protéines incluses dans le grain de pollen par la zone réceptrice du pistil. Malgré l’hermaphroditisme, le brassage génétique est optimal car la fécondation croisée est obligatoire.
.
Modalités de rencontre des sexes :
Les grains de pollen émis par les étamines ont besoin d’être transportés jusqu’aux organes femelles récepteurs afin que la fécondation puisse se faire. Bien qu’immobiles, les plantes utilisent des stratégies pour assurer les déplacements nécessaires :
.
Le transport par le vent caractérisait tous les gymnospermes (Pinus, Gingko, Juniperus …). Le pollen doit être particulièrement abondant (forme de gaspillage). C’est aussi le cas des arbres à chatons (ex.: Alnus glutinosa, Quercus …), qui ont des structures qui pendent dans le vent et recueillent les grains de pollen. Idem pour les graminées aux grandes étamines souples (ex.: Avena sat. …). On peut aussi citer le cas plus complexe de la Vallisnerie, plante aquatique : pour se rencontrer, les pieds femelles produisent une longue tige souple et une fleur qui arrive au ras de la surface, les pieds mâles libèrent des petits sacs de pollens qui montent en surface et dérivent jusqu’aux fleurs femelles qui dès que fécondées s’enroulent au fond pour mûrir (c’est-à-dire qu’on observe un retour à une vie aquatique chez un végétal aérien, un peu comme pour les dauphins, mammifères marins).
.
Le transport par les insectes s’est répandu et spécialisé avec l’évolution, au point que la vie animale et végétale sont souvent intriquées. Les fleurs s’adressent aux insectes par des appels liés à des contrastes de couleurs qui tranchent sur le vert ambiant et indiquent une direction à suivre (le centre en général) où se trouve le nectar, le pollen et les organes à féconder. La disposition des organes reproducteurs de la fleur est souvent particulièrement judicieuse par rapport à la zone nutritive (ex.: les labiées comme Lamium alb., Salvia off. …). Chez certaines plantes, on observe que la longueur du tube nectarifère correspond à la longueur de la trompe de tel papillon. Si un insecticide détruit ce papillon dans un secteur donné, la plante ne pourra plus se reproduire par graine.
.
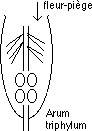
Le rapport à l’insecte est parfois cruel, ainsi Aristolochia clematitis et Arum triphylum dont les fleurs forment un tube coudé qui se termine par un renflement contenant les organes sexuels. Dans ce tube il y a des poils turgescents, raides et orientés vers le bas : les mouches qui rentrent dans la fleur passent sans encombre parmi les poils, mais elles ne peuvent ressortir ! Celles-ci se couvrent de pollen et dès que la fleur est fécondée, les poils se fanent et les insectes peuvent ressortir.
.
Les orchidées (ex.: la Vanille) ont établi avec les insectes des relations plutôt violentes : leurs fleurs sécrètent une substance odoriférante semblable à celle dégagée par les hormones sexuelles (phéromones) de certains insectes (guêpes …). Lorsque l’insecte mâle, attiré par l’odeur se rapproche de l’orchidée, il confond ses fleurs avec ses partenaires : tête, thorax, abdomen, poils, rayures de couleur, antennes, ailes, tout est parfaitement imité. Il se pose alors et cherche à copuler. Le pollen est concentré sur une seule étamine, aggloméré en deux petites masses, les pollinies. La fleur mâle possède un mécanisme qui, au moindre attouchement, catapulte les pollinies qui se collent sur la tête de l’insecte. Dépité devant le peu de réaction de cette femelle végétale, il s’en écarte et tente sa chance avec une autre femelle … qui n’est qu’une autre orchidée qu’il féconde. La perfection de ce système est confondante, d’autant que chaque espèce dégage un parfum qui n’attire qu’une seule catégorie d’insecte, et que l’émission de cette phéromone végétale est programmée quelques jours avant celle de l’insecte femelle, afin que les mâles ne soient pas enclins à leur préférer leurs partenaires naturelles.
.
Le transport par les oiseaux nectarivores (Colibris) a nécessité une adaptation de forme et de couleur : la couronne est réduite, les organes récepteurs de pollen sont hauts, les étamines un peu au dessous (ex.: la Passiflore, l’Avocatier).
.
Le transport par les chauves-souris intéresse des plantes (ex.: le Baobab, cf. image ci-dessous – sans doute un des plus gros et plus vieux arbre du monde !) à fleurs nocturnes, à nectar abondant et à odeur atroce (pour nous !).
.

.
Dès le début de l’ère tertiaire, le monde se couvre donc de plantes à fleurs dont se nourrissent des mammifères. Angiospermes et mammifères se diversifient, s’adaptent même à des milieux où la vie est très menacée. Ils s’y implantent et y créent des biotopes plus accueillants. Leur efficacité biologique, plus grande que celle de leurs prédécesseurs, leur permet d’aller plus loin dans la conquête de la planète. Il y a 50 millions d’années, le visage du monde ressemblait déjà à celui que nous connaissons. Ce visage résultait essentiellement du travail du monde végétal, poursuivi pendant plus de trois milliards d’années. C’est ainsi que fut peu à peu créée la biosphère, cette mince couche de sols arables, d’eau douce et d’air respirable dans laquelle prolifèrent les êtres vivants à la surface de la planète. Œuvre de la vie, elle est nécessaire à l’entretient de la vie.
.
Il y a 200 000 ans apparaissait l’homme…. Malgré les agressions constantes de notre civilisation envers son environnement, il existe encore sur notre planète plus de 300 000 espèces différentes de plantes. Ces végétaux, non centralisés, ni limités dans le temps (la reproduction végétative est presque toujours possible), s’offrent le luxe de « payer » les services qu’ils demandent au monde animal :
.
Si les liens entre le monde animal et le végétal sont évidents (ils arrivent même à produire les mêmes phéromones !), la sexualité animale paraît relativement monomorphe, comparée à celle du monde végétal qui a exploré toutes les solutions possibles d’une façon assez délirante.
.
Le médecin doit toujours prendre en compte le fait de cette évolution conjointe du monde végétal et du monde animal. Ce d’autant que « l’ontogénèse rappelle la phylogenèse« , c’est à dire que le développement embryonnaire humain récapitule toute l’histoire des espèces vivantes.
.
Nous étudierons successivement les quatre grands groupes d’angiospermes :
-> Les monocotylédones, de formule 3,
-> Les dicotylédones, de formule 5 (plus rarement de formule 4 ou 6)